线虫不喜欢被触摸。戳一下就会让蠕虫向后滑行,远离烦人的轻推。除非,也就是说,当它被戳时,蠕虫正在转动。 “不知何故,转动的行为使蠕虫改变了它对这种非常显着的刺激的反应方式,但我们不知道这种计算是如何在大脑中发挥作用的,”普林斯顿大学的神经科学家和研究人员Andrew Leifer说。西蒙斯全球大脑合作。
尽管对秀丽隐杆线虫大脑中的每个神经元和突触都有完整的连接组图,Leifer 和合作者仍然难以理解线虫的触摸和转动系统是如何相互作用的。 “当你开始追踪这些电路在哪里相遇时,你最终会进入这个密集的解剖连接网络,并且不清楚哪些路径最相关或信号如何通过它们,”Leifer 说。
随着时间的推移,团队发现他们探索的每条赛道都达到了类似的极限。 “我们最终意识到该领域的每个人都面临着同样的问题:连接组向你展示了假定的电路机制,但观察信息实际上是如何通过网络流动的却不那么简单,”Leifer 说。 “我们最终想到:为什么我们不只测量大脑中的所有功能连接,并提供一种可以类似于结构连接组使用方式的资源呢?”
在博士后Francesco Randi的带领下,该团队开展了一项大规模的工作,以光遗传学方式激活秀丽隐杆线虫大脑中的 302 个神经元中的每一个,同时对所有其他神经元的反应进行成像。由此产生的功能连接图谱于 8 月发布在 arXiv 上,描述了 10,000 多个神经元对之间的功能连接——其中许多是首次观察到的。
这些前所未见的连接之一恰好发生在来自转动和触摸电路的神经元之间。事实证明,一个神经元会抑制另一个神经元,即使它们之间没有直接的突触。相反,这种抑制是间接或“有效”连接的结果,在解剖图上是不可见的。 “在这个实验之前,我们甚至不知道这种联系的存在,现在它正在告知我们对运动协调的理解,”Leifer 说。 “我们现在很高兴看到其他电路计算可以通过功能连接组学来解释。”
这些实验是通过整合结构和功能连接测量来研究神经回路的日益增长的努力的一部分。 “功能连接”一词通常用于人类神经影像学,以描述跨大脑区域活动的相关性,但现代成像工具现在允许研究人员在单细胞分辨率下对模型生物进行类似的测量。在基于这一传统定义的研究中,相关的神经元活动表明功能相互作用,统计方法可以帮助估计哪个神经元正在影响哪个神经元。在一种更新的方法中,像莱弗这样的研究利用光遗传学的进步来更直接地探索神经元连接的方向性。通过激活单个细胞并成像其他细胞的反应,这些研究测量了神经元之间越来越多地被称为“因果连接”的东西。其他小组开始结合这些新的大规模解剖和活动数据集来测试关于大脑区域和行为的特定假设。尽管这项工作仍处于早期阶段,但这些发现突出了蠕虫、苍蝇和老鼠大脑中的重要网络特性,并阐明了电路结构和功能之间的复杂关系。
虫子的承诺
秀丽隐杆线虫通常被认为是了解结构如何产生功能的理想系统。 1986 年,它的连接组成为第一个被完全绘制出来的连接组,这是一项巨大的成就,为从感觉和运动到摄食和繁殖等蠕虫行为的无数发现提供了依据。尽管如此,接线图本身并不足以完全阐明动物更复杂的行为,这些行为往往依赖于更复杂的神经网络。在这里,广泛的功能工具提供了一个新的信息层,可以揭示神经回路以前隐藏的方面。 “这项技术终于赶上了蠕虫的承诺,”莱弗说。 “我们已经完成了几十年的解剖工作,但直到现在我们才能够在大脑规模和细胞分辨率上进行这些功能测量。”
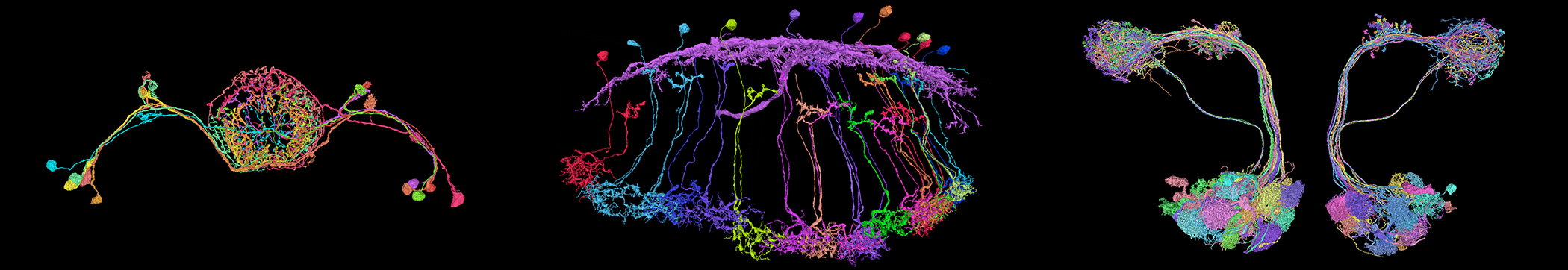
 Leifer 实验室的激活反应图显示了秀丽隐杆线虫神经元之间的功能连接。红色箭头表示兴奋性连接,而蓝色箭头表示抑制性连接。每个箭头的宽度反映了下游神经元反应的平均幅度,这是连接强度的一种度量。神经元的位置反映了它们的实际解剖排列。改编自 Randi 等人。 2022,arXiv。
Leifer 实验室的激活反应图显示了秀丽隐杆线虫神经元之间的功能连接。红色箭头表示兴奋性连接,而蓝色箭头表示抑制性连接。每个箭头的宽度反映了下游神经元反应的平均幅度,这是连接强度的一种度量。神经元的位置反映了它们的实际解剖排列。改编自 Randi 等人。 2022,arXiv。Leifer 和合作者制作的“扰动图”代表了一项重大的技术进步,它依赖于几种新技术。为了构建地图,研究人员必须刺激秀丽隐杆线虫大脑深处的单个神经元,同时高速成像所有其他神经元。这需要一个复杂的光学设置,将双光子刺激与单光子旋转圆盘共聚焦显微镜配对。它还需要一种非常规的光遗传学蛋白,这种蛋白可以被紫光激活,但不会对用于成像其他神经元钙反应的蓝光做出反应。 “我们花了很长时间寻找能够很好地结合在一起的蛋白质,”Leifer 说。最后,他们在一种新的多色蠕虫菌株中进行了所有这些实验,该菌株用不同的颜色荧光标记每个神经元,因此细胞更容易识别它们的功能活动何时注册到结构连接组。
这些实验中最引人注目的发现之一是在解剖图中无法检测到的一种功能连接形式:突触外通信。含有神经肽、单胺或其他神经递质的致密囊泡从突触外的部分神经元中释放出来,然后它们通过细胞外空间扩散到周围细胞上的受体上。虽然之前已经观察到这些信号,但预计它们不会在神经动力学中发挥重要作用。新发现表明,它们实际上在整个蠕虫大脑中相当普遍,并且对短期神经元活动有显着贡献,这表明它们的行为范围比以前想象的要广泛。
Leifer 说,这种“无线”信号只是功能连接不同于神经解剖学预测的一种方式。 “但我们也在了解哪些特征使结构和功能更加相似或更加不同。”例如,在显示较少突触外信号的蠕虫咽部神经元子网络中,研究人员发现电路结构和功能之间的一致性更好。
由 SCGB 研究员、维也纳大学的神经科学家Manuel Zimmer领导的第二项蠕虫研究使用了一种更经典的方法来描述秀丽隐杆线虫的功能连接。在这里,研究人员对蠕虫大脑的自发活动进行成像,然后测量每个神经元对之间的相关性。然后他们查看结构连接组是否可以预测功能连接模式。
结果于 2022 年 7 月发表在Current Biology上,揭示了令人惊讶的差异。化学突触并不是蠕虫功能连接的特别好的指标,这意味着两个直接连接的神经元不会同时始终活跃。电突触的存在,其中离子通过间隙连接在神经元之间流动,是一个更好的预测指标,但不是很多。考虑突触强度也无助于解剖连接组的预测能力。 “对于一对神经元来说,只有它们是否连接才重要;它们之间的突触数量没有区别,”齐默说。
事实上,每个神经元对的功能连接性主要是通过非局部电路特征来预测的,例如它们是否有共同的输入。 “大脑两侧没有解剖连接但接收相似输入的两个神经元通常会比直接相互连接的神经元表现出更多的相关活动,”齐默说。
这两项研究都强调了结合结构和功能信息的重要性,并提供了测量功能连接性的补充方法。自发活动可能更准确地反映神经元的正常行为方式,而系统地激活每个神经元可确保电路的任何部分都没有被探索。
飞行中的功能
与蠕虫大脑相比,果蝇大脑拥有大约 100,000 个神经元,为研究人员提供了更复杂的连接网络和更丰富的行为库可供探索。由于飞行连接组学的最新进展,他们现在可以在更广泛的范围和更精细的分辨率下进行研究。 Mala Murthy是普林斯顿大学的神经科学家和 SCGB 研究员,是推动全脑连接组学发展的科学界的一员。她的最新项目是与Sebastian Seung的一项名为FlyWire的合作,它允许神经科学家众包,从而加快校对神经重建的过程,这是完成连接组的最后一步(更多信息,请参阅“连接的连接组”)。 Murthy 及其同事现在正在利用这些全面的重建来探索神经动力学,重点关注感觉回路的结构如何产生神经元的特征选择性。
Murthy 实验室使用 FlyWire 绘制了跨越苍蝇大脑两个半球的 500 多个神经元之间的解剖连接图。由此产生的接线图能够预测神经元的听觉反应特性。 Amy Sterling 的动画以 FlyWire 中绘制的听觉神经元为特色,由 Baker 等人出版,Current Biology 2022
其中一项研究于 2022 年 7 月发表在Current Biology上,重点关注雌蝇的听觉回路。该电路的主要功能是帮助协调对男性求爱歌曲的适当响应,该歌曲在两种主要模式之间交替 – 正弦和脉冲。先前的工作表明,听觉电路可能被设计为将正弦信号和脉冲信号分离成两个独立的处理流。
为了验证这一假设,博士后Christa Baker首先确定了每个听觉神经元是否喜欢正弦或脉冲歌曲,然后将结果与 FlyWire 图中神经元的解剖连接进行比较。虽然一些神经元确实对正弦或脉冲歌曲有明显的偏好,但研究小组惊讶地发现,许多神经元表现出混合或中等偏好。所有这些不同的神经元实际上并没有被分隔成不同的正弦和脉冲通路,而是紧密地相互连接,它们之间没有明显的层次结构。研究人员尚不确切知道为什么以这种方式设计电路,但他们的模型表明,这种互连有助于产生混合偏好人群的调整。此外,电路内的重复可能有助于雌蝇在更长的时间尺度上处理求爱歌曲的特征,或将信息与其他感觉信号整合。
“很明显,苍蝇的听觉系统不仅仅是一个简单的前馈感觉回路,而是包含许多循环连接,以及与其他大脑区域的互连,”Murthy 说。 “我认为这让人们感到惊讶,因为到目前为止,我们一直在前馈框架内研究这些感觉通路,但苍蝇大脑更像是一个多感觉反馈控制系统,新的结构和功能数据确实清楚地说明了这一点。 ”
功能连接也已通过飞行中的扰动方法进行测量,尽管不像 Leifer 的蠕虫研究那样在全脑范围内进行测量。在 2018 年发表在eLife上的一篇论文中,Janelia 研究园区的Vivek Jayaraman实验室同时使用光遗传学刺激和钙成像来测试果蝇大脑中 70 个细胞对之间的连通性。将这些因果连接测量与解剖连接组进行比较,Jayarman、 Romain Franconville和合作者发现功能连接似乎比预期的要稀疏得多——换句话说,激活神经元在下游神经元中产生的反应比基于连接组预期的要少得多。
然而,Jayaraman 指出,他们的光遗传学协议旨在提供最小的刺激,目的是只激活直接的突触连接。这意味着在这些实验条件下可能无法检测到较弱突触或需要多个突触前神经元共激活的额外功能连接。
尽管如此,Jayaraman 说:“很明显,解剖结构只是用于计算的支架,然后将功能电路放置在上面。”作为一名训练有素的工程师,Jayaraman 经常依靠计算模型来探索这种神经网络动力学。 “我们长期以来一直试图通过理论将结构与功能联系起来,现在我们可以通过功能连接组学直接测试很多这些想法。”
目前尚不清楚从蠕虫或苍蝇中学到的具体经验将如何转化为其他生物,但这些早期发现表明,每个物种可能采用不同的机制来优化其神经回路的处理能力。 “结构和功能连接组之间的匹配将因不同物种而异,”Murthy 说。 “突触外连接可能有助于秀丽隐杆线虫的大脑与更少的神经元建立更大的交流库,但神经系统越大,你就越能将这种特异性与更多的细胞联系起来。” Murthy 和 Jayaraman 认为,最有可能在无脊椎动物和脊椎动物物种中推广的是电路拓扑结构,而不是拓扑结构。物理图和机制可能不同,但网络的结果功能和计算可能是相似的。 “问题是:复杂的连通性是如何产生这些人口信号和行为的?”穆尔蒂说。 “我们现在只是触及表面,但这是未来几年我们将拥有的乐趣。”
绘制小鼠大脑图
无脊椎动物模型,它们的小大脑和稀疏的电路,为结构连接组学铺平了道路。但是在测量功能连接性方面,小鼠实验室也取得了长足的进步。在 SCGB 研究员Karel Svoboda位于 Janelia 研究园区的实验室的一项新研究中,现就职于特拉维夫大学的Arseny Finkelstein基于最近的技术进步来测量小鼠运动皮层的因果连接性。通过单个神经元的高速激活和成像,Finkelstein 能够在 30 分钟的时间内绘制出 50 万对神经元之间的连接——这是之前小鼠研究的 10 倍放大。
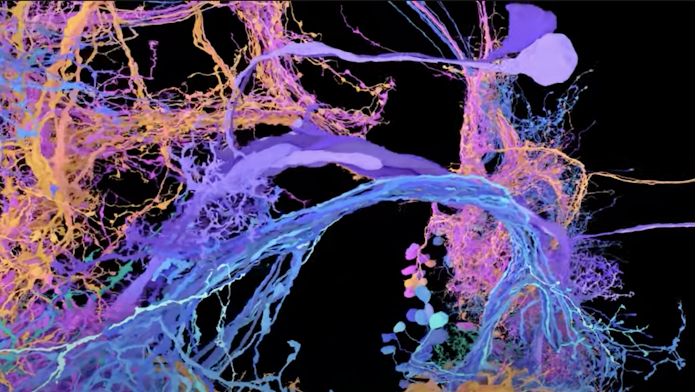
 在 Svoboda 实验室的实验中,单个运动神经元被光遗传学光刺激(粉红色),而其他兴奋性神经元(绿色)的反应是用双光子钙成像测量的。显示出显着活动反应(红色)的细胞被认为在功能上与受刺激细胞相关。阿尔塞尼·芬克尔斯坦
在 Svoboda 实验室的实验中,单个运动神经元被光遗传学光刺激(粉红色),而其他兴奋性神经元(绿色)的反应是用双光子钙成像测量的。显示出显着活动反应(红色)的细胞被认为在功能上与受刺激细胞相关。阿尔塞尼·芬克尔斯坦使用这种策略,芬克尔斯坦探索了未经训练的运动皮层的功能组织。他说:“我们对运动皮层的了解大部分来自受过专门训练的动物,但在学习发生之前这些电路是什么样子的?”为了回答这个问题,芬克尔斯坦专注于前外侧运动皮层 (ALM),这是一个参与协调鼠标舔动作的区域。他首先让幼稚的老鼠执行一项简单的舔舐任务,以确定每个神经元偏好的舔舐方向。然后他刺激每个神经元,同时测量其他神经元的反应。
3 月份在里斯本举行的 Cosyne 会议上公布的结果揭示了 ALM 的高水平职能组织。类似的调谐神经元似乎被分组在皮质迷你柱中,它们相互兴奋,但在柱外抑制不同调谐的神经元。这个主题已经在灵长类动物的初级感觉区域和某种程度上在果蝇的大脑中看到过,但在这些高阶额叶皮质中不太确定。
另一个主要发现是存在中枢神经元、具有密集传出连接的细胞、较弱的特征选择性和更可靠的放电模式。研究人员尚不清楚这些中枢神经元在做什么,但他们推测这些细胞被放置在整个电路中以帮助组织全脑活动。 “我们认为,这些中枢神经元可能会启动运动序列并协调更广泛的皮层动力学,而不是代表运动计划的特定特征,”Svoboda 说。
Zimmer 的C. elegans研究还确定了中枢神经元,突出了用于协调全脑活动模式的潜在保守电路基序。在蠕虫中,与这些中枢神经元之一共享连接是神经元之间功能连接的主要预测指标。事实上,抑制这些中枢神经元中的一些就足以破坏蠕虫的全脑相关性。随着功能连接性研究继续揭示每个模型系统中的网络特性,科学家们有兴趣了解其他哪些电路基序在物种之间是保守的或专门用于每个模型系统。
最大的悬而未决的问题之一——也是最具挑战性的答案——是这些连接组在每个生物体中跨个体和跨时间尺度的稳定性如何。随着成像、重建和校对连接组的方法变得更加精简,苍蝇和蠕虫研究人员可能会率先解决这个问题。例如,Murthy 对比较单个果蝇的结构和功能很感兴趣。 “果蝇大脑的相对刻板印象对神经科学家来说是一个巨大的优势,但可能仍然存在一些有趣的可变性来源,这些来源可能会令人兴奋地理解并与功能相关。”
脊椎动物更大、更复杂的大脑可能会更加多变。 “哺乳动物系统专门用于学习和灵活性,因此我们可以期待看到它们的结构和功能连接以及旨在支持这种灵活性的电路图案有更多的可变性,”Svoboda 说,他最近从 Janelia 搬到他现在正在西雅图的艾伦研究所研究全脑连接。他估计我们距离拥有完整的小鼠连接组至少还有十年的时间,尽管一些小鼠研究人员开始在功能成像后收集较小脑容量的回顾性重建。
研究人员最终希望了解功能连接性如何不仅从物种到物种或从个体到个体,而且在学习和大脑状态之间发生变化。这就是芬克尔斯坦选择在幼稚、未经训练的老鼠身上进行功能连接测量的原因。 “拥有’基线’功能连接的快照将使我有一天可以询问有关如何修改电路的问题,”他说。 “但是在去那里的路上还有很多问题需要回答。”